Edafología. Volumen 5. Diciembre 1998. pag 29-39.
SALINITY - FERTILITY INTERACTIONS ON EARLY GROWTH OF MAIZE (Zea mays L.) AND NUTRIENT UPTAKE
Pablo Zalba and Norman Peinemann
Departamento de Agronomía, Universidad Nacional del Sur 8000 Bahía Blanca, Argentina.
Plants are generally classified as being either halophytes or glycophytes based mainly on their ability to grow in saline environments. According to Flowers et al. (1986) the most useful criteria for separating these two groups is the ability of halophytes to complete their life cycle at excess concentrations of 100 to 200 mM NaCl. In contrast, glycophytes are plant species characterised by relatively low salt tolerance (Miller and Doescher, 1995). Halophytes typically achieve high salt tolerance by inclusion of salts while glycophytes exhibit relatively limited salt tolerance by excluding salt from leaves cells. Glycophytes may osmotically adjust by producing biochemically-compatible organic solutes within the cytoplasm. Notwithstanding, in many cases an important factor in salt damage is dehydration due to the extracellular accumulation of salt as suggested in the Oertli hypothesis (Flowers et al., 1991). It is well known that the response of glycophytes to salinity may be influenced by the plant species and by environmental conditions, and also varies at different stages of development within the same species. For instance, maize has a relatively high salt tolerance during the seed germination stage, but is less tolerant of salinity during the later stages of growth (Ayres & Hayward; 1948). Furthermore, the apparent salt tolerance of many agricultural crops varies with the soil fertility level i.e. several crops showing exceptionally high apparent salt tolerance at a low fertility level, become more sensitive when adequately fertilised. However, some data have been recorded about a real increase in salinity tolerance under improved fertility conditions (Feigin, 1985). On the other hand, in the case of an extremely salt tolerant species like Lavatera arborea L., which is physiologically adapted to saline environments, an increase in the level of soil fertility (mineral nutrients) resulted in an increase in growth (Okusanya, 1980).
Owing to the fact that ionic absorption always increases together with the salt concentration in irrigation water and soil solution, nutritional requirements of cultivated plants should be satisfied with greater difficulty. A large body of literature has been written on the effects of salinity on the nutrition with N, P and K by glycophytes from which the following information was selected:
Nitrogen: There is ample evidence of root absorption competition between Cl- and NO3- by plants [ De Wit et al. (1963) for rice; Weigel et al. (1973) for beans; Berstein et al. (1974) for some vegetable crops; Kafkafi et al. (1982) for tomato]. However, there is no evidence to show that increasing amounts of nitrogen under saline conditions avoid such effect of competition.
Phosphorus: The effect of salinity on phosphate concentrations in the plant is variable. In some cases negative effects were reported [Ravikovitch & Yoles (1971); Champagnol (1979)]. On the contrary, Cerda & Bingham (1978) reported a yield increase in tomato owing to P application at all salinity levels studied (Kafkafi, 1984). There is no evidence with regard to competition for uptake of H2PO4- and Cl- by plants. Furthermore, because under saline conditions root growth is limited, probably greater amounts of P are needed to supply the total demand.
Potassium: Potassium contents in plant tissue progressively decrease with the increase in salinity owing to a higher absorption of sodium. Such effect was observed by Bernstein et al. (1974) and Finck (1976) in wheat; Kaddah & Ghowail (1964) in maize; Bierhuizen & Ploegman (1967) and Rush & Epstein (1976) in tomato; Allison (1964) in beans and some varieties of carrot; Patel (1973) in sorghum, maize, wheat and Sudan grass; Cheng (1984) in sunflower. The Na/K ratio is also an important feature in determining the sensitivity of protein synthesis to salt. High Na/K ratios inhibit protein synthesis through the decline in translational efficiency (Wyn Jones et al., 1979). However, if an additional surplus of K is necessary to maintain a constant Na/K ratio in plants is not known.
Finally, an important subject that must be mentioned is the case of some glycophytes which are sensitive to salinity only during specific short periods of their life cycles. Evidences for this hypothesis were reported by Bernstein & Hayward (1958) and Lunin et al. (1963) among others, several decades ago. For instance, Kaddah & Ghowail (1964) and Pasternak et al. (1985) showed that the most sensitive period of maize to salinity is during the first three weeks after sowing. Alleviation of salinity stress during this sensitive period resulted in a significant increase in the salt resistance of this crop. In this regard, the purpose of this study was to evaluate the effect of fertiliser elements (N - P - K) on nutrient uptake of maize plants at different water salinity levels during the post-emergence period. In addition, the effect of salt concentration on seed germination was determined.
1. Effect of salinity on germination of maize seeds: Fifty maize seeds (cv DEKALB 3F21) treated with the fungicide "Vitavax" were placed in Petri dishes lined with four layers of Whatman N° 44 filter paper. Seven solutions of NaCl were prepared to obtain electrical conductivities of 0, 3, 6, 9, 12, 16 and 20 dS/m respectively. Each solution was poured into four dishes totalizing 200 seeds and four replicates per treatment. Later, Petri dishes were kept in an incubator at 25°C for ten days. Seeds were considered germinated when radicles measured 5 mm. To estimate the rapidity of germination the method proposed by Tucker & Wright (1965) was adopted. The index of rapidity of germination (I) is obtained from the equation I = (N/2 - a)/b where "a" is the intercept and "b" the slope of the adjusted line of the germination curve and N/2 = 50. In this method index values are directly expressed in number of days to achieve 50 % germination (Tucker & Wright, 1965).
2. Plant studies with nitrogen, phosphorus and potassium under salinity conditions: Laboratory studies were carried out with maize plants to determine the effect of salinity and fertility on dry matter production and nutrient uptake. Dry soil (120 g) from the zone of Bahía Blanca, Argentina (sandy loam, Typic Haplustoll) was weighed in 4 cm deep plastic pots with a diameter of 8 cm. Forty maize seeds (cv DEKALB 3F21) treated with the fungicide "Vitavax" were placed in each pot. Two replicates per treatment were made. Pots were then placed in a growth chamber at 12 hours artificial light/darkness intervals at 25oC and 15oC respectively. Pots were weighed every 1 or 2 days and watered to maintain soil moisture between -0.033 and -0.05 MPa. The principle of the experiment is based on the exhaustion of soil available nutrients through an excessive density of plants in a short time (21 days). It is assumed that available nutrients have been exhausted after three weeks, therefore, maximum dry matter production has been obtained under these controlled conditions. Consequently, it is not necessary to consider the total number of plants at the end of the trial. Another advantage of this experiment consisted in using seed with high hybrid vigour (cv DEKALB 3F21) which greatly reduced experimental variability.
Effects of nitrogen, phosphorus and potassium applications under salinity were studied in separate experiments.
Nitrogen: Amonium nitrate was used as nitrogen source at rates equivalent to 0, 25 and 50 mg N kg-1 at six salinity levels (Water EC: 0, 3, 6, 9, 12, 16 and 20 dS/m). Water EC were obtained using standard solutions of NaCl. The trial was considered finished after 21 days; therefore dry matter and fresh matter production were then determined. Finally, 0.5 g of triturated sample was placed in a furnace at 550°C for two hours. Ashes were solubilised in 0.1N HNO3 to determine chloride volumetrically by the Mohr method. NO3-N was determined on the original triturated sample using an improved extraction procedure adapted for ultraviolet spectrophotometry (Heanes, 1982).
Phosphorus: Sodium phosphate (NaH2PO4.H2O) was used as a phosphorus fertiliser at rates equivalent to 0, 20 and 50 mg P kg-1. Seven salinity levels were studied (Water EC: 0, 3, 6, 9, 12, 16 and 20 dS/m).
Fresh and dry matter production were evaluated after 21 days. Sodium, potassium and calcium were determined by flame spectrophotometry, phosphorus by the Murphy-Riley colorimetric method and chlorides by the Mohr method.
Potassium: KCl rates equivalent to 0, 40 and 80 mg K kg-1 were studied at six salinity levels (Water EC: 0, 3, 6, 9, 12 and 16 dS/m). Fresh and dry mater production were evaluated after 21 days. Sodium, potassium and chlorides were determined in the ashes with the same methodology mentioned above.
3. Root experiments: Saline solutions were prepared with KNO3 (5 mmol), KH2PO4 (1 mmol) and NaCl (0, 6, 18, 42 and 90 mmol) to obtain electrical conductivities of 0.6, 1.2, 2.4, 4.8 and 9.6 dS/m respectively. Young ten-days-old excised maize roots (cv DEKALB 3F21) were placed into the saline solutions for 30 minutes to favour the flow of ions into the root cells. Roots were then washed with distilled water and placed in 0.1N H2SO4 for another 30 minutes to promote the movement of ions outwards. Three replicates per salinity level were made. Roots were dried to constant weight at 50°C. Nitrate, phosphorus potassium, sodium and chloride were determined using this methodology.
Statistical Analysis
Data obtained on the plant growth parameters were subjected to analysis of variance and comparison of means was made according to Tukey's test (Steel and Torrie, 1980).
1. Salinity effects on germination: maize showed a relatively high tolerance to salinity during the germination stage as can be seen in Fig. 1. This observation is in total agreement with results obtained by Ayers & Hayward (1948), Maas et al. (1983), Pasternak et al. (1985), among others. Rapidity of germination in salinity levels 0, 3 and 6 dS/m was I=2 i.e. 50% germination at the second day, while germination energy (GE) decreased from 97.5% to 79% respectively. At salinity level 20 dS/m, 50% germination was achieved at the sixth day (I=6) and GE fell to 2.5% (Table 1). Notwithstanding the toleration of salinity observed during this stage, both rapidity of germination and GE were statistically reduced in salinity levels 12, 16 and 20 dS/m respectively (P<0.05). Of greater relevance, however, is the fact that no statistically significant differentiation was found between salinity levels 0, 3 and 6 dS/m (P<0.05). It is also remarkable that shapes of germination curves (Fig. 1) are very similar to that obtained by Pasternak et al. (1985) with cv KWS 752 corn seed. Therefore, such different hybrid materials as DEKALB 3F21 and KWS 752, do not introduce a great variability with regard to salt tolerance during germination. Nevertheless, Schubert & Lauchli (1986, 1990) revealed strong differences in the root surface of two maize cultivars in relation to sodium permeability of the plasmalemma which may represent different strategies to tolerate salinity within the same species.
2. Plant experiments with nitrogen, phosphorus and potassium.
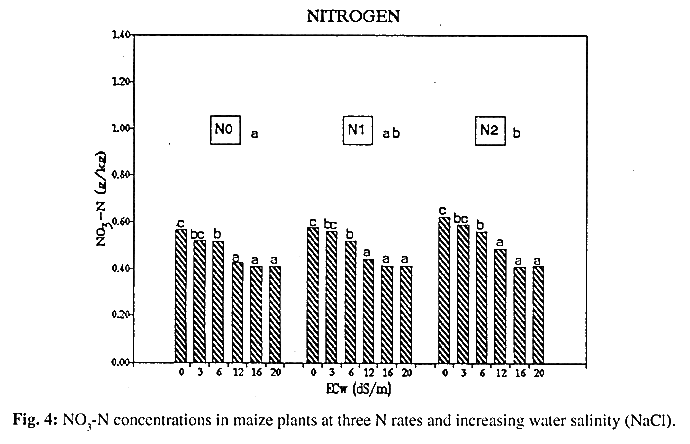
Nitrogen: Fresh matter (FM) and dry matter (DM) production were both significantly reduced (P<0.01) at the following salinity levels: 12, 16 and 20 dS/m. In addition, increasing nitrogen availability at these three levels did not lead to increase FM production significantly (P<0.05) (Fig. 2, Fig. 3). In accordance with the latter, Selassie & Wagenet (1984) showed that maize growth was not improved as a result of applying N. Moreover, no beneficial effect was exerted on salinity tolerance of young maize plants by nitrogen applications. NO3-N concentration in the shoot tended to diminish with increasing levels of salinity in spite of a nitrogen surplus. Furthermore, a high negative correlation ( r = -0.84**) between NO3- and Cl- was observed (Fig. 4, Fig. 5). According to these observations the interaction process between both anions seems to be competitive, at least during the early growth stage of maize.
In the case of tomato, 15N concentration in lowly salt- stressed plants was higher than that in the controls at all three uptake times studied i.e. 7-, 14- and 21-days harvest, but it was lower in highly stressed plants except in 21-days harvest (Pessarakly & Tucker, 1988). On the other hand, 15N concentration in cotton plants was statistically higher for moderately stressed than for lowly stressed plants (Pessarakly & Tucker, 1985). These results indicated that in both cases plants have somewhat adjusted to salinity and its effects on N uptake. Such differences with regard to nitrogen uptake under NaCl stress reflect variations in salt tolerance of these cultivated plants i.e. cotton > tomato > maize.
Phosphorus and potassium: Application of high rates of P and K did not consistently increase maize production of FM and DM at all the levels of salinity studied (Fig. 2). H2PO4-P concentrations in plant were initially lower at salinity level 3 dS/m but rapidly increased reaching 0.3 g/kg d. wt at higher salinities. This difference was found to be statistically significant at the 0.01 probability level, as can be seen in Fig. 6. It seems to be that there was not a clear competitive interaction process involved between H2PO4- and Cl- during the early growth stage of maize. An increase in levels of salinity resulted in a decrease in growth but DM production could not be improved as a result of applying P or K. The Na/K ratio in plants increased proportionally with the increasing salt concentration indicating that greater amounts of K in the media did not lead to meliorate this ratio in plants. On the contrary, Na absorption increased independently of K concentration (Figs. 6, Fig. 7). According to Bernstein et al. (1974), high rates of P aggravated salt injury in maize and, also, increasing K concentration in the media from 0.4 to 2 me/l had little effect on leaf K contents and yield.
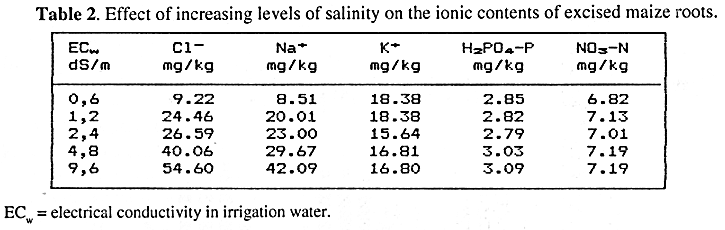
Results obtained from root experiments indicated that Na+ and Cl- uptake increased together with salinity similarly to what was observed in shoots. However, N, P and K contents remained constant (Table 2). Owing primarily to different absorption conditions, higher concentrations of NO3-N and H2PO4-P were observed in roots. These results support the idea that competition between NO3- and Cl- should be localised in leaves or shoot cells. Part of the reason for this assumption is the fact that NO3-N contents in roots remained constant at all the levels of salinity studied. Fuqua et al. (1974) arrived to a similar conclusion with regard to a burley tobacco species.
Fresh and dry matter production of maize plants were both significantly reduced by increasing salt concentration (NaCl) in irrigation water. Greater availability of nutrient elements such as N, P and K did could not counteract the ionic absorption effect, moreover, the response of young maize plants to nutrient applications tended to diminish with increasing salinity. A High negative correlation (r = -0.8488) between NO3- and Cl- concentrations in shoots was determined. However, phosphorus and potassium concentrations remained constant.
As a complement, results from root experiments indicated that the competitive interaction processes between NO3- and Cl- recorded in shoots was not observed at root level. Thus, it is proposed that the competition process between both anions must be regulated at specific sites in the leaves cells rather than on the root surface.
Finally, it was demonstrated again that maize is relatively tolerant to salinity during the germination stage and this behaviour is considered to be crop specific.
Allison L. E. 1964. Salinity in relation to irrigation. Adv. Agron. 16, 139.
Ayers A. D. and Hayward H. E. 1948. A method for measuring the effects of soil salinity on seed germination with observations on several crop plant. Soil Sci. Soc. Am. Proc. 19, 224.
Bernstein L., Francois L. E. and Clark R. A. 1974. Interactive effects of salinity and fertility on yields of grains and vegetables. Agron. J. 66, 412.
Bernstein L. and Haward H. E. 1958. Physiology of salt tolerance. Ann. Rev. Plant Phys. 9, 25.
Bierhuizen J. F. and Ploegman C. 1967. Salt tolerance of tomato. Meded. Dir. Tuinbouw Neth. 30, 302.
Cerda A. and Bingham F. T. 1978. Yield, mineral composition, and salt tolerance of tomato and wheat as affected by NaCl and phosphorus nutrition. Agrochimica 22, 140.
Champagnol F. 1979. Relationships between phosphate nutrition of plants and salts toxicity. Phosphorus Agric. 76, 35.
Cheng S. F. 1984. Effects of salinity, fertility and water on sunflower yield and nutrient uptake. Diss. Abstr. Intern. B (Sciences and Engineering) 44, 3600.
De Wit C. T., Dijkshoorn W. and Noggle J. C. 1963. Ionic balance and growth of plants. Versl. Landbouwkd Onderz. 69, 15.
Feigin A. 1985. Fertilisation management of crops irrigated with saline water. Plant and Soil 89, 285.
Finck A. 1976. Soil salinity and plant nutritional status. Proc. Intern. Conf. Manag. Saline Water for Irrigation. Texas Tech. Univ. 199.
Flowers T. J., Hajibagheri M.A. and Yeo A.R. 1990. Ion accumulation in the cell walls of rice plants growing under saline conditions: evidence of Oertli hypothesis. Plant, Cell and Environment 14, 319-325.
Fuqua B. D., Leggett J. E. and Sims J. L. 1974. Accumulation of nitrate and chloride by burley tobacco. Can. J. Plant Sci. 54, 167-174.
Kaddah M. A. and Ghowail S. L. 1964. Salinity effect on the growth of corn at different stages of development. Agron. J. 56, 214.
Kafkafi U. 1984. Plant nutrition under saline conditions. In Soil Salinity under Irrigation. I. Shainberg and J. Shalhevet Eds. Springer Verlag, Berlin, 349 p.
Kafkafi U., Valoras, N. and Letey J. 1982. Chloride interaction with nitrate and phosphate nutrition in tomato. J. Plant Nutr. 5, 1369.
Lunin J. Gallatin M. H. and Batchelder A. R. 1963. Saline irrigation of several vegetable crops at various growth stages. I. Effect on Yield. Agron. J. 55, 107.
Maas E. V., Hoffman G. J., Chaba G. D., Poss J.A. and Shanon M.C. 1983. Salt sensitivity of corn at various stages. Irrig. Sci. 4, 45.
Miller R. F. and Doescher P. S. 1995. Plant Adaptations to Saline Environments. In Wildland Plants: Physiological Ecology and Developmental Morphology. Donald J. Bedunah & Ronald E. Sosebee Eds. Denver, Colorado. p. 440-478.
Okusanya, O. T. 1980. The effect of salinity and nutrient level on the growth of Lavatera arborea sea water micro- organism. Oikos 35, 49.
Pasternak D., DeMalach Y. and Borovic I. 1985. Irrigation with brackish water and to alternating brackish-fresh-brackish water irrigation. Agric. Water Manag. 10, 47.
Patel P. M. 1973. Salinity-fertility interactions for five different crops in relation to yield and chemical composition. Diss. Abstr. Intern. 34, 20.
Pessarakli M. and Tucker T. C. 1985. Uptake of nitrogen-15 by cotton under salt stress. Soil Sci. Soc. Am. J. 49, 149.
Pessarakli M. and Tucker T. C. 1988. Dry matter yield and nitrogen-15 uptake under sodium chloride stress. Soil Sci. Soc. Am. J. 52, 698.
Rush D. W. and Epstein E. 1981. Comparative studies on the sodium, potassium and chloride relations of a wild halophytic and a domestic salt-sensitive tomato species. Plant Phys. 68, 1308.
Selassie T. G. and Wagenet R. J. 1981. Interactive effect of soil salinity, fertility and irrigation interval on field corn. Irrig. Sci. 2, 67.
Steel, R. G. D. and Torrie J. H 1980. Principles and procedures of statistics: A biometrical approach. 2nd ed. McGraw-Hill, New York.
Tucker H. and Wright N. L. 1965. Estimating rapidity of germination. Crop Sci. 5, 398.
Weigel R. C., Schillinger J. A., Mc Caw B. A., Gauch H. G. and Hsiao E. 1973. Nutrient-nitrate levels and accumulation of chloride in leaves of snapbeans and root of soybeans. Crop Sci. 13, 411.
Página Principal | Presentación EDAFOLOGIA | Indice vol. 5 | Principio página
{kind=link}
{kind=link}